
תַקצִיר
ב-50 השנים האחרונות הקדשתי את הקריירה שלי לחֵקֶר שאלות יסודיות בביולוגיה, במיוחד בתחום הגנטיקה. השאלות האלה נוגעות לחלק מהתהליכים הבסיסיים ביותר של החיים, כמו למשל כיצד תא מתפקד כיחידה עצמאית ועם זה מְתַקְשֵׁר עם סביבתו? וגם, כיצד התא משתמש במידע הגנטי שלו, שמאוחסן בדנ''א (DNA), ומעביר אותו לדורות התאים הבאים? במאמר הזה אני רוצה לקחת אתכם למסע בחלק מעבודתי כחוקר. אתאר לכם את הממצאים העיקריים שלי על דנ''א ועל השעתוק שלו לרנ''א שליח (mRNA) דרך מנגנון מורכב שנקרא רנ''א פּוֺלימֶרַאז II. הרנ''א שליח הזה בסופו של דבר מתורגם לחלבונים הממלאים מגוון תפקידים מרכזיים בתוך היצור החי, כולל בניית תאים, תגובה לאותות סביבתיים, זירוז תגובות כימיות והעברת אותות בין רקמות רחוקות. לבסוף, אספר לכם על חלק מהשאלות הפתוחות המרתקות שאנו עובדים עליהן בימים אלה, ואסיים עם טיפים אישיים עבורכם – מדעני העתיד.
פרופסור רוג'ר קורנברג זכה בפרס נובל בכימיה לשנת 2006 עבור מחקריו על הבסיס המולקולרי של שעתוק אאוקריוטי.
דַּרְכִּי אל הרנ''א
אבי היה ביוכימאי, והוא זכה בפרס נובל בשנת 1959 על עבודתו בחֵקֶר תהליכי שכפול דנ''א. אבא שלי חלק את ההתלהבות המחקרית האדירה שלו עם כל מי שהיה מוכן להקשיב, ואני ספגתי את אהבתו של אבי למדע. למדתי מתמטיקה, פיזיקה וכימיה בקולג', ובדוקטורט שלי חקרתי את קרום התא – המֶּמְבְּרָנָה (איור 1 משמאל). לקרום התא יש את התפקיד החשוב ביותר בחיים, מאחר שמעטפת כזו שמקיפה את התא החי היא מה שמגדיר את הקיום של התא – היחידה הבסיסית של כל יצור חי.
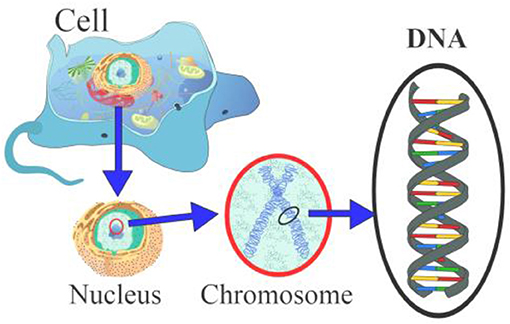
- איור 1 - קרום התא והגרעין שמכיל את הכרומוזום ואת הדנ''א.
- התא הוא היחידה הבסיסית של החיים. גרעין התא, שהוא מרכז המידע של התא, מכיל כרומוזומים. כל כרומוזום הוא מבנה בצורת 'X' (מוקף באדום) שבו חלק מהדנ''א. האיור הותאם מ: Wikipedia (https://en.wikipedia.org/wiki/Chromosome).
לקראת סוף עבודתי על קרום התא ידעתי כי ברצוני להמשיך לחקור את הפיזיקה והכימיה שקשורים למדעי החיים. באותו זמן, התחום של ביולוגיה מולקולרית, שעוסק בין השאר בחֵקֶר המרכיבים של התא והתהליכים שמתרחשים בו, התפתח במהרה. פיתוחים טכנולוגיים חדשים אפשרו לפענח את המבנה של חלבונים פשוטים. אז התוודעתי לבעיה של מבנה הכרוֹמוזוֹם (ראו איור 1). הכרומוזום הוא מבנה שבו החומר הגנטי, הדנ''א, מאורגן בתוך גרעין התא. מבנה הכרומוזום סיקרן ביולוגים מבניים בגלל חשיבותו הבסיסית של הדנ''א והמבנה הפשוט לכאורה שלו. באותה עת כבר הכרנו את אבני הבניין של הכרומוזום; הוא מורכב מדנ''א ומארבעה חלבונים קטנים מאוד. כל מה שהיינו צריכים להבין היה כיצד הדנ''א וארבעת החלבונים האלה מאורגנים יחד ויוצרים את מבנה הכרומוזום.
התברר שהבעיה הזו לא הייתה פשוטה. בסופו של דבר פתרתי את הבעיה באמצעות מיון של כמה מאמרים רלוונטיים מִבֵּין מאות רבות של מאמרים שנכתבו על מבנה הכרומוזום. המאמרים האלה סיפקו לי את הרמז הראשוני לפתרון. ביצעתי ניסויים רלוונטיים ובאמצעותם הצלחתי לחבר יחד את חתיכות הפאזל ולפתור את הבעיה של מבנה הכרומוזום. מבנה זה הוכח מאוחר יותר כנכון באמצעות שיטה הקרויה קריסטלוגרפיה בקרני רנטגן.
באופן טבעי, אחרי פתרון הבעיה הזו רציתי לגלות מהן ההשלכות של מבנה הכרומוזום על ביולוגיה, על החיים עצמם. כלומר, מדוע הדנ''א מאורגן באופן המסוים הזה בתוך הכרומוזום לשם ביטוי של מידע גנטי. ביטוי של גֶּן מתחיל בתהליך שנקרא שִׁעְתּוּק, שבו נוצרת מולקולת רנ''א שליח (mRNA) מדנ''א. רנ''א שליח הוא מולקולה דומה לדנ''א, אך בעלת מבנה ותפקוד שונים. רנ''א שליח בנוי מסליל יחיד קצר שנבנה לפי רצף הדנ''א, ובעל הרכּב מעט שונה מהדנ''א. בניגוד לדנ''א שבנוי משני סלילים (איור 1, מימין), מולקולת הרנ''א שליח בנויה מסליל יחיד קצר, שמהווה עותק של מִקְטָע דנ''א מסוים. הרנ''א מהווה חוליה מקשרת בין המידע הגנטי שבדנ''א לחלבונים שנוצרים לפי מידע זה. לכן, כדי לחקור את תפקיד הכרומוזום ולהבין את ביטויוֺ של מידע גנטי, התחלתי עם אחד משלושת האנזימים שמבצעים שעתוק. האנזים הזה נקרא רנ''א פּוֺלימֶרַאז II [1].
מדנ''א לרנ''א – מנגנון השעתוק של רנ''א פולימראז II
תפקידו של מנגנון השעתוק של הרנ''א פולימראז II הוא ליצור רנ''א שליח. כפי שהזכרתי קודם, רנ''א שליח הוא החוליה המקשרת בין קוד הדנ''א והחלבונים המסוימים שמיוצרים ממנו. מנגנון השעתוק של רנ''א פולימראז II מכיל כמעט 60 סוגים של חלבונים! אתאר את שלושת הרכיבים העיקריים [2]: אנזים הרנ''א פולימראז II, חלבונים שנקראים גורמי שעתוק כלליים וקומפלקס של קבוצות שונות של חלבונים שנקרא מתווך.
אנזים רנ''א פולימראז II – מבנה ותפקוד
אנזים הרנ''א פולימראז II (איור 2A) הוא המבנה שבתוכו מתרחש תהליך השעתוק. משמעות הדבר היא שדנ''א נכנס אל האנזים הזה מכיוון אחד, ותוצר הרנ''א שליח יוצא מהצד השני. הרבה מהעבודה שלנו הייתה סביב פתרון המבנה המורכב של האנזים הזה באמצעות מדידות רנטגן. לאחר פענוח המבנה של האנזים הזה לבדו, הצלחנו גם לפענח את המבנה שלו במהלך תהליך השעתוק. כלומר, כשגם הדנ''א וגם הרנ''א נוכחים בתוכו במהלך תהליך השעתוק הפעיל (איור 2B).
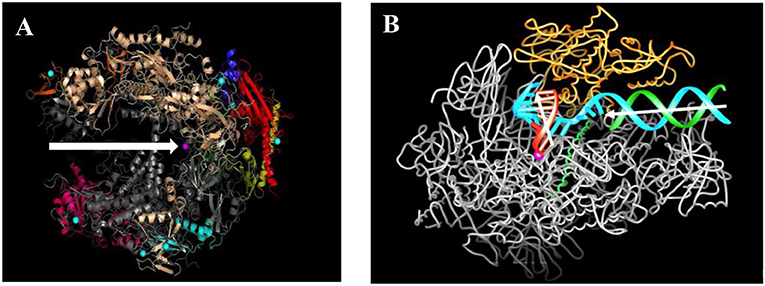
- איור 2 - מבנה אנזים הרנ''א פולימראז II לפני השעתוק ובמהלכו.
- (A) אנזים הרנ''א פולימראז II מורכב מ-12 תת-יחידות (שמיוצגות בצבעים שונים) וכמה עשרות אלפי אטומים. יש לו תעלה מרכזית (חץ לבן) שמובילה ליון מגנזיום (נקודה ורודה). האזור שבו נמצא יון המגנזיום נקרא ''אתר פעיל'' מאחר ששם הרנ''א השליח נבנה מהדנ''א. (B) סליל דנ''א כפול (טורקיז וירוק) נכנס דרך התעלה המרכזית של אנזים הרנ''א פולימראז II (חץ לבן אופקי) ומתפצל קרוב למרכז האנזים. סליל דנ''א אחד (טורקיז) מסובב ב-90 מעלות למעלה ליד האתר הפעיל (חץ לבן שפונה למעלה). לפי התבנית של סליל הדנ''א הזה נבנה סליל קצר של רנ''א שליח (סליל אדום קצר במרכז). הקומפלקס המשולב הזה של דנ''א ורנ''א שליח יוצא מהאנזים בכיוון מאונך לכיוון שממנו הדנ''א נכנס במקור. קרדיט לאיור: פרופ' רוג'ר קורנברג.
הרנ''א פולימראז II מורכב מ-12 תתי-יחידות שונות, שמוצגות בצבעים שונים באיור 2A, וכמעט 30,000 אטומים. לרנ''א פולימראז II יש תעלה מרכזית, שמובילה ליון מגנזיום. התעלה המרכזית היא המקום שבו מתרחש השעתוק. סליל כפול של דנ''א נכנס לתעלה המרכזית ושני הסלילים נפרדים (איור 2B). סליל אחד מכוֹפָף ליד יון המגנזיום במרכז האנזים. במיקום זה, שנקרא אתר פעיל, הרנ''א השליח נבנה על פי ההוראות שבסליל הדנ''א המכופף. לבסוף, מבנה משולב של דנ''א-רנ''א שליח יוצא מהאנזים בזווית של כ-90 מעלות יחסית לזווית שבה הדנ''א נכנס תחילה לאנזים.
גורמי שעתוק כלליים – רכיבי מפתח נוספים בביטוי גֶּנִים
השלב החשוב ביותר בתהליך השעתוק הוא האִתחוּל שלו. כאשר דנ''א משועתק לרנ''א שליח, הוא לא משועתק במלואו. רק חלק מסוים מאוד בדנ''א משועתק. החלק הזה נקרא גֶּן. כל גן מכיל מידע על ייצור של חלבון מסוים בגופנו. הרנ''א פולימראז II זקוק לחמש מולקולות נוספות כדי לזהות גן ספציפי ולשעתק אותו. המולקולות האלה הן חלבונים שנקראים גורמי שעתוק כלליים, והם באים במגע עם אנזים הרנ''א פולימראז II במהלך תהליך השעתוק (ראו עיגולים אפורים באיור 3 למטה). באופן כללי, אתם יכולים לחשוב על גורמי השעתוק הכלליים האלה כרכיבים במנגנון השעתוק שמסייעים ''להדליק'' או ''לכבות'' גנים מסוימים.
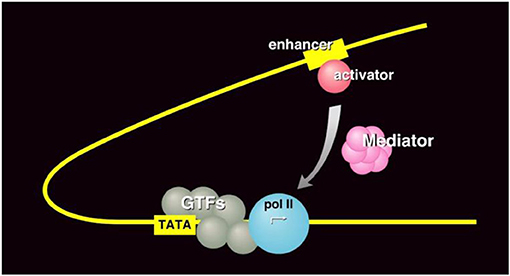
- איור 3 - מנגנון השעתוק של רנ''א פולימראז II.
- למטה: גורמי שעתוק כלליים (GTFs, באפור) מְתַקְשְׁרִִים עם אנזים הרנ''א פולימראז II (pol II, בכחול) כדי לאתחל את שעתוק הדנ''א בתוך האנזים. המתווך (בוורוד) משמש כחוליה מקשרת שמספקת לאנזים הרנ''א פולימראז II מידע על בקרת גנים מגורמים שונים בתוך התא ומחוץ לו. במקרה הזה, המתווך מעביר מידע מחלבון מפעיל (אדום) על הפעלה של גן מסוים לשעתוק. קרדיט לאיור: פרופ' רוג'ר קורנברג.
כפי שראינו קודם לכן, כאשר הדנ''א עובר בתוך הרנ''א פולימראז II הוא צריך להתכופף כדי להיות משועתק לרנ''א שליח. דנ''א בצורתו הרגילה (סליל כפול) הוא קשיח מאוד, ולא יכול להתכופף. כדי לכופף אותו הוא צריך להיפתח לשני סלילים נפרדים, ואז הוא הופך גמיש לחלוטין ויכול להתכופף בחופשיות לכל הכיוונים. כאן גורמי השעתוק הכלליים נכנסים לתמונה: אחרי שגורמי השעתוק הכלליים מזהים את תחילתו של הגן במולקולת הדנ''א הישרה, הם פותחים את הדנ''א ומכופפים אותו ליד האתר הפעיל של השעתוק ברנ''א פולימראז II. באופן הזה, גורמי השעתוק הכלליים מאתחלים את תהליך השעתוק.
מתווך – ''איש הביניים'' של בקרת גנים
בתהליך של שעתוק דנ''א צריכות להתקבל החלטות חשובות מאוד: אילו גנים לשעתק, באיזה מקום בגוף ומתי. קבוצת ההחלטות האלה נקראת 'בקרת ביטוי גנים', והיא קריטית עבור התפקוד התקין של גופנו. המתווך הוא קבוצה של חלבונים שגילינו ב-1990 [3], והוא רכיב מרכזי במנגנון בקרת הגנים: מעבד את כל המידע הגנטי שמגיע מתוך התא ומחוץ לתא, ומעבירו לרנ''א פולימראז כדי לשלוט בהחלטה אם לשעתק גן מסוים.
איור 3 מראה באופן סְכֶמָתִי את תפקודו של המתווך בתהליך השעתוק: המתווך (בוורוד) מְקַשֵּׁר בין חלבון שנקרא מפעיל (באדום), אשר משפיע על ''הדלקה'' של שעתוק גן, לבין אנזים הרנ''א פולימראז II (בכחול). במילים אחרות, המתווך משמש כ''איש ביניים'', שמספק לאנזים הרנ''א פולימראז מידע שקשור לבקרת ביטוי גנים.
שאלות פתוחות לעתיד
אני רוצה לגרות את סקרנותכם ולספר לכם על שני נושאים פתוחים שקשורים למה שסיפרתי לכם במאמר הזה. הנושאים האלה הם חוד החנית של מחקר עכשווי בביוכימיה, ואנו עובדים על שניהם במעבדה שלי ממש בימים אלה.
הנושא הראשון קשור למבנה של הכרומוזום. בשלב מסוים בחלוקת התא, הדנ''א בכרומוזום מכווץ באורכו פי כ-10,000, כך שהחומר התורשתי עצמו, שקודם לכן תפס את כל גרעין התא, נדחס לגבולות המצומצמים של הכרומוזום. מה שאנו יודעים בוודאות על מבנה הכרומוזום (שציינתי בתחילת המאמר) מסביר את התקצרות אורך הדנ''א רק פי חמישה אולם לא פי 10,000. כלומר, השאלה הפתוחה היא: כיצד הדנ''א בכרומוזום נדחס עוד פי 2,000 ממה שאנו יודעים להסביר?
הנושא השני קשור במתווך שדנו בו קודם ובבקרה של ביטוי גנים. כפי שראיתם באיור 3, אנו יודעים שהמתווך מספק מידע של בקרת גנים לאנזים הרנ''א פולימראז II. אולם כיצד מידע הבקרה מעובד על-ידי המתווך? כיצד בדיוק המידע המעובד הזה מועבר אל הפולימראז? כיצד המתווך מסייע בקיפול של דנ''א מחוץ למבנה הכרומוזום כדי לשעתק אותו? יש לנו רעיונות על מה שעשוי להתרחש ואיך, אולם אלה עדיין חידות פתוחות שאנו מנסים לפתור.
המלצות למוחות צעירים
במובנים רבים, השאלות שציינתי כאן הן אותן שאלות בסיסיות שאיתן התחלתי את המסע האקדמי שלי. כפי שאתם ודאי יודעים, שאלות מדעיות רבות הן מורכבות ונדרשות שנים רבות של מחקר כדי לענות עליהן. מדע הוא מאתגר, הוא דורש עבודה מאומצת שיכולה לעיתים להיות קשה ולעיתים מתסכלת. אולם עבורי התגמולים המזדמנים שווים לגמרי את המאמץ. אם אתם אוהבים מדע ורוצים לעסוק בקריירה מדעית, העצה הראשונה שלי אליכם היא ליהנות מהפעילות עצמה, ללמוד כיצד ליהנות מהדברים הקטנים שמרכיבים את פעילויות היומיום. במקרה שלי, לדוגמה, הפעילויות האלה הן ניסוייות – ערבוב והמסה של חומרים שונים ויצירת תמיסות עבור הניסויים שלי. אני באופן אישי אוהב כל אחד מהצעדים הקטנים האלה, ואני אוהב לבלות במעבדה.
היבט חשוב נוסף הוא ללמוד כיצד לחוות כישלון כמשהו ממריץ ומאתגר – כהזמנה לנסות שוב עם אותה מידת ציפייה להצלחה כמו בפעם הקודמת. מדי פעם משהו טוב קורה, ואז מדענים טובים צריכים לא להאמין מייד לתוצאות. ראשית, עליכם לוודא שלא מדובר בטעות, לכן אתם צריכים לחשוב על דרכים להוכיח שאתם טועים. מדענים טובים חושבים על דרכים מתוחכמות ביותר להוכיח שהם טועים. אם הם לא מצליחים להוכיח שטעו, אפילו באמצעות ניסויים משוכללים, אז ככל הנראה הם גילו משהו. רגעי הגילוי האלה הם רגעים ייחודיים מאוד ובלתי נשכחים בקריירה של מדענים, ובעיניי הם מצדיקים לחלוטין את המאמצים שהושקעו בתהליך.
מילון מונחים
דנ''א חומצה דיאוקסיריבונוקלאית, (DNA): ↑ החומר שנושא מידע גנטי באורגניזמים חיים. הדנ''א בנוי משני סלילים שיוצרים סליל כפול והוא ממוקם בגרעין התא.
חלבונים (Proteins): ↑ מולקולות גדולות הנמצאות בכל התאים החיים, אשר ממלאות תפקידים חשובים רבים ביצורים חיים.
כרומוזום (Chromosome): ↑ המבנה הבסיסי שבו דנ''א מאורגן בתוך גרעין התא.
גן (Gene): ↑ יחידה תפקודית בסיסית של דנ''א, המכילה את המידע ליצירת חלבון ביצורים חיים.
שעתוק (Transcription): ↑ השלב הראשון בביטוי גן, אשר בו נבנית מולקולת רנ''א על פי הרצף של מקטע דנ''א מסוים.
רנ''א שליח (mRNA): ↑ אחד מסוגי הרנ''א בתא החי אשר מעורב בתהליך של ייצור חלבונים על פי המידע בדנ''א.
אנזים (Enzyme): ↑ מולקולה ביולוגית גדולה שמזרזת תגובות כימיות המתרחשות בגוף או בתוך התא.
גורמי שעתוק כלליים (General transcription factors): ↑ קבוצה של חמישה חלבונים המסייעים באתחול של תהליך שעתוק הדנ''א לרנ''א שליח.
מתווך (Mediator): ↑ קומפלקס רב-חלבוני המעבד מידע על בקרת ביטוי גנים, ומעביר מידע זה לגורמי שעתוק כלליים ולרנ''א פולימראז II.
הצהרת ניגוד אינטרסים
המחבּר מצהיר כי המחקר נערך בהיעדר כל קשר מסחרי או כלכלי שיכול להתפרשׁ כניגוד אינטרסים פוטנציאלי.
תודות
ברצוני להודות לנועה שגב, בוגרת התוכנית לאנרגיה ע''ש ננסי וסטיבן גרנד (GTEP), טכניון, ישראל, על עריכת הריאיון שהיווה את הבסיס למאמר זה, ועל כתיבה משותפת של המאמר.
מקורות
[1] ↑ Cramer, P., Bushnell, D. A., and Kornberg, R. D. 2001. Structural basis of transcription: RNA polymerase II at 2.8 Å resolution. Science 292:1863–76. doi: 10.1126/science.1059493
[2] ↑ Boeger, H., Bushnell, D. A., Davis R., Griesenbeck, J., Lorch, Y., Strattan, J. S., et al. 2005. Structural basis of eukaryotic gene transcription. FEBS Lett. 579:899–903. doi: 10.1016/j.febslet.2004.11.027
[3] ↑ Kelleher, R. J., Flanagan, P. M., and Kornberg, R. D. 1990. A novel mediator between activator proteins and the RNA polymerase II transcription apparatus. Cell 61:1209–15. doi: 10.1016/0092-8674(90)90685-8