
摘要
正如人类拥有不同的语言, 鸟类也各有独特的鸣唱。 这些鸣叫声帮助鸟类相互识别并寻找配偶。鸟鸣的演化方向大概取决于两个因素: 一是栖息地, 二是身体特征。首先, 栖息地会影响鸣叫声的传播距离;其次, 体型大小和喙的形状会限制鸟类能发出的声音类型。我们以吸蜜鸟为研究对象, 探索了这些因素如何影响鸟鸣演化。研究发现, 体型是解释鸟鸣差异的首要因素——体型更大的鸟类会发出更低频的鸣叫, 就像体型高大的人的嗓音更低沉!部分研究表明, 栖息地也会影响鸟鸣演化, 但这很可能源于不同栖息地鸟类的体型差异。这些发现具有重要意义: 随着未来气候变化, 鸟类体型预计将变小, 它们的交流方式也有可能随之改变!
鸟鸣为何重要?
鸟鸣是动物界最广为人知的声音之一, 堪称大自然的"歌单"。你是否曾在清晨被鸟鸣唤醒, 并好奇它们为何歌唱?正如我们能通过声音识人, 鸟类也能通过鸣声辨识同类。例如, 雄鸟通过鸣唱吸引雌鸟, 鸣声帮助它们识别同一物种并选择配偶。全球现有超过 5, 000 种鸣禽, 这意味着存在极其丰富的鸟鸣声!科学家认为, 鸣声差异促进了新种鸣禽的演化。研究演化能帮助我们了解过去引发物种变迁的因素, 从而了解当今现存物种, 并推测它们如何适应未来的变化。
哪些因素可能影响鸟鸣演化?
鸟鸣是一种复杂的性状, 科学家认为其演化方向取决于栖息地和身体特征(名为形态学)。由于雄鸟通过鸣唱吸引雌鸟, 雌鸟能否听见雄鸟的歌声至关重要。鸟类生存的环境会影响其鸣声——可参阅 Frontiers for Young Minds 上这篇 关于城市噪音如何影响鸟鸣的文章。森林或茂密环境会吸收声音, 限制鸟鸣声的传播距离;相比之下, 沙漠或草原等开阔地区树木和吸音物体较少, 鸟鸣声能传播得更远。由于低频率声音比高频率声音的传播距离更远, 可以推测: 在茂密环境中, 经过长期演化的鸟类会发出比开阔环境中的鸟类频率更低的鸣叫声, 以确保被同类听见(图 1) [1]。
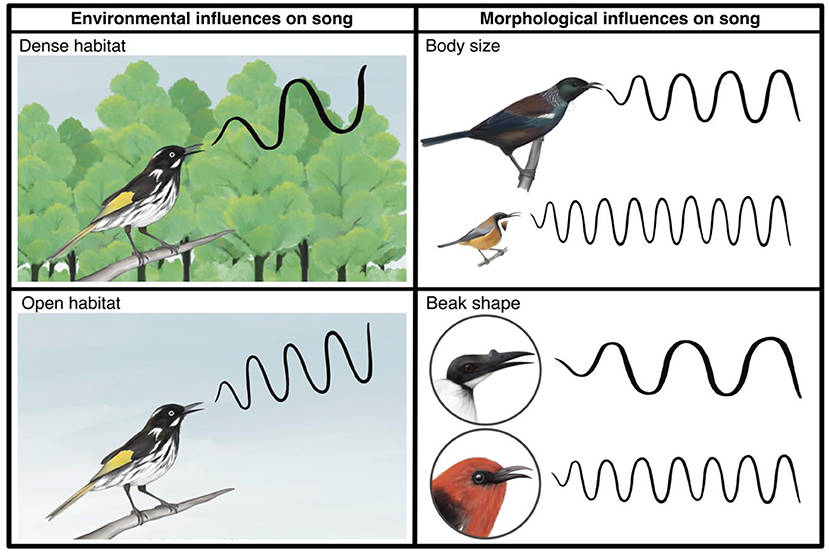
- 图 1 - 环境与形态对鸟类鸣声的预测影响示意图。
- (左图)环境对鸣声的影响: 生活在茂密栖息地的鸟类预计比开阔地区的鸟类产生更低频的鸣声。(右图)形态对鸣声的影响: 体型较大的鸟类预计比较小的鸟类产生更低频的鸣声;此外, 喙较大的鸟类预计比喙较小的鸟类鸣唱频率更低。声波图形表示频率: 较宽的波代表低频鸣声, 较窄的波代表高频鸣声(插图: Eleanor Hay)。
研究鸟类体型大小及结构特征的"形态学"也被认为会影响鸟鸣演化。特别是, 体型大小以及喙的尺寸与形状预计在其中起着关键作用: 鸟类体型会影响其发声频率, 这是因为体型决定鸣管大小。鸣管是鸟类的发声器官, 类似人类的喉部。鸣管尺寸影响声音频率 [2], 因此体型更大的鸟类预计会发出更低频的声音(图 1)。喙的尺寸与形状同样影响鸣声: 如同小号或长笛等乐器, 鸟喙的形状与大小决定了气流与声音振动模式, 从而影响鸣声频率。更大的乐器/鸟喙预计会产生更低频的声音(图 1)。此外, 喙的尺寸和形状会限制其开合灵活性, 进而影响发声。较大、较长或较弯曲的鸟喙无法像短小、纤细且平直的鸟喙那样快速开合, 因而被认为会影响鸣唱节奏 [3]。
显然, 栖息地与形态学能以多种方式影响鸟鸣。为弄清这些影响, 必须考虑这些因素之间的内在关联。形态学与栖息地并非相互独立。事实上, 鸟类体型和鸟喙形态与其生存环境密切相关。例如, 寒冷环境中生活的鸟类往往体型更大以保持体温, 而温暖环境中生活的鸟类可能体型较小以利于散热。喙的大小和形状也取决于鸟类食性: 长而弯曲的喙通常用于吸食花蜜, 短而直的喙则适合捕食昆虫或啄食种子。更复杂的是, 体型也会影响喙形: 大型鸟类往往拥有更大的喙。以往的研究通常只关注栖息地或形态学对鸟鸣的影响, 但这些因素其实是相互关联的, 我们需要综合考量栖息地与形态学的共同影响, 才能真正理解鸟鸣的演化机制。
吸蜜鸟
我们以吸蜜鸟为研究对象, 探究了栖息地与形态学对鸟鸣演化的共同影响。吸蜜鸟是一个包含 190 多个物种的鸟类类群, 分布于澳大利亚、新几内亚、新西兰及南太平洋地区。所有吸蜜鸟都源自同一祖先, 因主要取食植物花蜜而得名, 但部分吸蜜鸟也食用果实和昆虫。与其他地区体型娇小的蜂鸟和太阳鸟等食蜜鸟类不同, 澳大利亚最大的吸蜜鸟——黄垂蜜鸟——比最大的太阳鸟还要重三倍以上!吸蜜鸟在可能影响鸣声的特征上展现出丰富的多样性: 体重从不足 10 克到超过 350 克不等;鸟喙形态各异, 既有捕食昆虫的短直喙, 也有吸食花蜜的长弯喙;栖息地涵盖从干旱沙漠到湿润热带的各种环境(图 2)。这种多样性使吸蜜鸟成为探索鸟鸣演化的理想研究对象。

- 图 2 - 吸蜜鸟的形态多样性展示。
- 图中物种依次为: (A) 歌吸蜜鸟, (B) 黄吸蜜鸟, (C) 噪吸蜜鸟, (D) 白额澳䳭, (E) 白耳吸蜜鸟, (F) 新荷兰吸蜜鸟, (G) 紫颊吸蜜鸟, (H) 黑额矿吸蜜鸟, (I) 暗吸蜜鸟(供图: Eleanor Hay)。
我们如何探索鸟鸣演化?
为了对吸蜜鸟的鸣声进行分类, 我们使用了在线数据库中的雄鸟鸣叫录音, 共分析了来自 163 种吸蜜鸟的 550 余段录音。针对每段录音, 我们生成了声谱图。这种图像能像乐谱一样按时间轴显示声音的音高变化。我们从每段鸣声中提取了三项参数: 峰值频率、频带宽度和鸣唱节奏(图 3)。频率以赫兹(Hz)为单位(以物理学家海因里希 ⋅ 赫兹命名), 峰值频率即鸣声中最响亮或最突出的音, 对声音传播至关重要, 因为最可能被潜在配偶听到。频带宽度是整个鸣声中最高与最低音的频率差值, 反映物种能发出的音域范围。鸣唱节奏则通过单位时间内的发音数量计算, 体现鸟类的鸣唱速度(图 3)。这些参数共同描绘了跨物种的鸣声多样性。
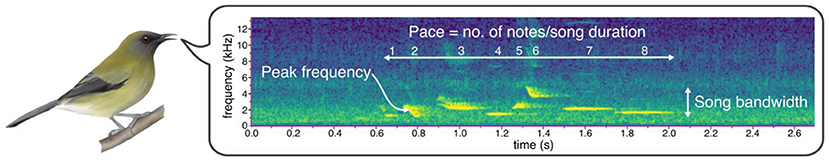
- 图 3 - 新西兰钟雀的鸣唱声谱图, 横轴为时间(秒), 纵轴为频率(千赫兹), 图中标注了本研究所用的三项鸣声参数。
探索吸蜜鸟的鸣声演化时, 必须考虑物种间的系统发育关系。系统发育树类似家族谱系图, 能揭示物种亲缘关系及其演化历程。幸运的是, 我们通过前期研究已掌握吸蜜鸟的系统发育关系 [4]。系统发育关系不容忽视, 因为亲缘较近的物种通常比远缘物种具有更相似的性状— — 例如, 无论栖息地与形态是否相似, 近缘鸟种的鸣声往往更接近。这种系统发育效应就像子女在外貌上通常更像父母而非他人。
体型更大的鸟类鸣声频率更低
研究发现, 推进吸蜜鸟鸣声演化的主要因素是体型。体型更大的鸟类能发出更低频的鸣声, 且鸣唱节奏更慢。这是因为体型决定鸣管大小。较小的鸟类通常拥有更小的鸣管, 其振动速度更快, 因而能产生更高频的鸣声;而较大鸟类的鸣管更大, 通常以较低频率振动, 发出的声音也更低沉。这是动物界的普遍规律: 体型影响着哺乳动物、昆虫、鱼类、爬行动物和两栖动物的交流方式 [5]。实际上, 体型更大的人类也往往拥有更浑厚的嗓音!尽管我们之前推测鸟喙形态会影响鸣声演化, 但研究未发现相关证据。虽然鸟喙形态与体型相关, 但在吸蜜鸟中, 体型与鸣声演化的关联更为显著, 喙形未单独表现出影响。
环境因素有何影响?
研究确实发现, 环境对吸蜜鸟的鸣声具有一定影响: 在茂密生境中, 节奏更慢、频率更低、频带更宽的鸣声更占优势。这初步表明栖息地可能参与推动鸟鸣演化。然而, 我们也发现, 茂密生境中的吸蜜鸟体型普遍大于开阔生境。最终分析显示, 栖息地类型对鸣声的影响更可能源于不同生境中的体型差异。这进一步证实体型是鸟鸣演化的主要驱动因素。以往的研究仅发现微弱证据表明环境能影响其他鸟类的鸣声, 但学界认为环境对动物交流的影响可能更广泛存在于青蛙、哺乳动物等其他类群 [1]。因此, 我们总体上认为栖息地主要通过改变物种体型等性状间接影响鸣声演化。
未来鸟鸣可能会如何演化?
既然已经了解鸟鸣的演化历程, 我们可预测其未来变化趋势。气候变化是物种面临的最大威胁之一。已有研究表明, 气候变化正导致鸟类体型缩小, 以便更高效地散热 [6]。根据体型决定鸣声频率这一发现, 未来更小的体型很可能导致更高频的鸣声。气候变化还将导致栖息地萎缩与生态转型, 而研究证实栖息地会影响体型等性状, 从而参与塑造鸣声。因此, 气候变化与栖息地破坏将对鸟鸣演化产生深远影响, 改变动物的交流方式, 继而重塑大自然的"歌单"。
术语表
物种 (Species): ↑ 能够自然繁殖并产生可育后代的一群生物个体。
进化 (Evolution): ↑ 种群或物种特征随时间发生的变化, 通常是为了更好地适应生存环境。
性状 (Trait): ↑ 生物个体或物种的特定特征, 可能由遗传因素和/或环境因素决定。
形态学 (Morphology): ↑ 生物体各部分的尺寸、形状及排列方式。
频率 (Frequency): ↑ 每秒产生的声波数量(以赫兹计量), 影响声音音调的高低。
鸣管 (Syrinx): ↑ 鸟类的发声器官。
声谱图 (Spectrogram): ↑ 音频信号的可视化呈现, 展示频率随时间的变化, 类似于乐谱。
系统发育(或系统发育树) (Phylogenetic (or Phylogeny)): ↑ 系统发育学研究生物类群间的演化历史。系统发育树是展示这些亲缘关系的示意图, 类似家族谱系图。
利益冲突声明
作者声明本研究不涉及任何潜在商业或财务关系。
致谢
谨此感谢 Steven Chown、Matt McGee 和 Craig White 对本文所依据的原创研究做出的重要贡献。
AI 人工智能工具使用声明
本文中所有图表附带的替代文本(alt text)均由 Frontiers 出版社在人工智能支持下生成。我们已采取合理措施确保其准确性,包括在可行情况下经由作者审核。如发现任何问题,请随时联系我们。
原文
↑Hay, E. M.、 McGee, M. D.、 White, C. R., 和 Chown, S. L. 2024. 体型决定吸蜜鸟的鸣声. Proc. R. Soc. B. 291:20240339. doi: 10.1098/rspb.2024.0339
参考文献
[1] ↑ Ey, E., and Fischer, J. 2009. The “acoustic adaptation hypothesis” – A review of the evidence from birds, anurans and mammals. Bioacoustics. 19:21–48. doi: 10.1080/09524622.2009.9753613
[2] ↑ Ryan, M. J., and Brenowitz, E. A. 1985. The role of body size, phylogeny, and ambient noise in the evolution of bird song. Am. Nat. 126:87–100. doi: 10.1086/284398
[3] ↑ Podos, J. 2001. Correlated evolution of morphology and vocal signal structure in Darwin’s finches. Nature. 409:185–188. doi: 10.1038/35051570
[4] ↑ Hay, E. M., McGee, M. D., and Chown, S. L. 2022. Geographic range size and speciation in honeyeaters. BMC Ecol. Evol. 22:1–14. doi: 10.1186/s12862-022-02041-6
[5] ↑ Gillooly, J. F., and Ophir, A. G. 2010. The energetic basis of acoustic communication. Proc. R. Soc. B. 277:1325–1331. doi: 10.1098/rspb.2009.2134
[6] ↑ Gardner, J. L., Peters, A., Kearney, M. R., Joseph, L., and Heinsohn, R. 2011. Declining body size: a third universal response to warming? Trends Ecol. Evol. 26:285–291. doi: 10.1016/j.tree.2011.03.005